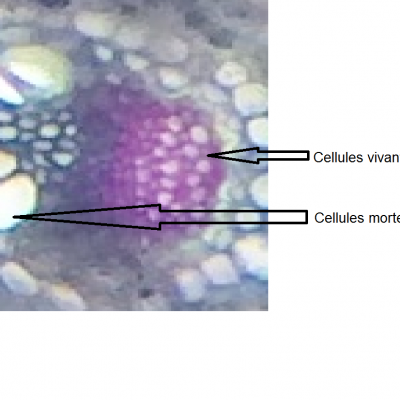
La plante domestiquée
Thème 2 Enjeux planétaires contemporains
La plante domestiquée
Les plantes sont des organismes autotrophes et, à ce titre, occupent une position essentielle dans l'alimentation de l'Humanité. Leur utilisation par l'être humain est une très longue histoire qui commence par la cueillette, se développe avec la culture et se poursuit actuellement par l'utilisation de la génétique et des technologies de pointe. La maîtrise de l'exploitation des plantes constitue donc un enjeu majeur pour l'humanité, que ce soit pour l'alimentation, la santé ou le développement durable.
- La domestication des plantes sauvages : L'exemple du maïs
Le maïs est cultivé aujourd’hui dans presque toutes les régions du monde, c’est la troisième céréale du monde, il est la base de l’alimentation d’une grande partie de la population mondiale. Le maïs actuel est le fruit d'une longue sélection des plants présentant le plus d'intérêt pour l'être humain, parfois même au détriment de la plante elle-même.
- La sélection empirique
Dans un premier temps, cette sélection des plants présentant un intérêt pour l'être humain a été empirique, c'est à dire non raisonnée et en se basant uniquement sur des caractéristiques évidentes et visibles.

On peut d'ailleurs résumer les critères de sélection par l'homme en comparant le téosinthe et le maïs
| Elements comparés | Plante | Epis femelles | Glumes | Grains | Nombre de rangées de grains |
| Téosinte | Une tige principale portant au sommet un panicule, mais des ramifications latérales longues portant au sommet des panicules et sur des ramifications secondaires des épis femelles |
Pratiquement pas de rachis, grains soudés entre eux qui se désarticulent à maturité peu de grains (8 à 10) Epi : 4 fois moins long |
Glumes indurées qui entourent le grain : protection | Simple | Deux rangées de grains |
| Maïs | Une tige principale qui porte au sommet un panicule. Des ramifications latérales très courtes portant des spadices |
Rachis important, qui ne se désarticule pas Grand nombre de grains (jusqu’à 500) Epi 4 fois plus long |
Glumes souples qui n’entourent pas le grain | Par paire, 2 fois plus longs | Plusieurs rangées autour du rafle |
L'être humain a donc progressivement sélectionné les plants:
Possédant des ramifications latérales courtes (moins encombrants, on peut en mettre plus dans le même espace)
Dont les épis femelles étaient les plus longs, qui portaient les grains les plus gros et les plus nombreux (plus d'apport nutritif pour une seule plante)
Dont les grains ne se désarticulaient pas (plus simple pour la récolte)
Dont les glumes étaient réduites (moins de temps de préparation)
Seuls ces grains étaient utilisés pour être replantés.
Ces caractéristiques, si elles sont très utiles pour la consommation et ne posent pas de réels problèmes dans un agrosystème, se révèlent en revanche nettement moins favorables à la plante dans son écosystème naturel. Les grains qui ne se désarticulent plus ne peuvent tomber et germer, rendant la reproduction impossible, la disparition de la glume entraîne une plus forte perte par prédation. Ainsi, le maïs actuel est incapable de se reproduire sans l'action humaine.
| Type d'habitat et sélection | Ecosystème naturel peu modifié | Agrosysteme |
|
sélection |
||
| Naturelle exercée sur l'ancêtre sauvage | Exercée par l'Homme sur la plante cultivée | |
| Caractéristiques génétiques | Conséquences sur les phenotypes | |
| Architecture de la plante |
Forme buissonnante avec ramifications très développées: sensibilisation réduite aux vents. Une tige principale portant au sommet un |
Forme peu ramifiée et plus haute favorable à une mise en culture dense (forte compétition); Inflorescence pas trop haute pour ne pas être sensible aux vents. Une tige principale qui porte au sommet un panicule (inflorescence mâle). Des ramifications latérales très courtes portant les inflorescences femelles. |
| Floraison | Absences de synchronisation : limite les effets de facteurs climatiques (amérique centrale absence d’hiver) | Floraison sur le même pied mais décalé: évite l'autopollinisation (favorise la fécondation croisée) mais synchronisme dans la floraison ce qui facilite la récolte : les grains sont mûrs tous en même temps. Mais dépend du climat. |
| Nombres de graines-Forme épis femelles |
Pratiquement pas de rachis, grains soudés entre eux qui se désarticulent à maturité ; peu de grains (8 à 10) Epi 4 fois moins long |
Rachis important,qui ne se désarticule pas ; grand nombre de grains (jusqu’à 500) Epi 4 fois plus long |
| Enveloppes protectrices de la graine à maturité |
Glumes indurées qui entourent le grain : protection Epaisses : protection contre les facteurs environnementaux défavorables |
Glumes souples qui n’entourent pas le grain. Réduction voire disparition. |
| Taille des fruits et des graines-réserves | Taille et réserve réduites. Simple et 2 rangées de grains |
Tailles plus importantes, réserves en biomolécules importantes. Par paire, 2 fois plus longs et 4 à 5 fois plus lourds Plusieurs rangées de grains autour du rafle La plante cultivée produit un grand nombre de grains, plus gros et plus longs donc qui contiennent plus de réserves : la domestication a permis une augmentation de rendement. |
| Devenir des graines à maturités | Libération par le fruit : dispersion favorisée. Les grains de Téosinte tombent sur le sol, ils sont mangés mais la cupule résiste aux enzymes digestives, ce qui permet leur dissémination. |
Pas de libération : récolte facilitée A maturité ces grains ne tombent plus car l’épi ne peut pas se désarticuler, ils ne possèdent pas de cupule solide donc la récolte est complète (sans perte) et plus rapide. De ce fait la dissémination des grains est impossible, la plante ne peut donc plus se reproduire à l’état sauvage. |
Légende : en vert, caractéristique favorable à la plante mais défavorable à l'être humain ; en rouge, caractéristique favorable à l'être humain mais défavorable à la plante ; en bleu, caractéristique favorable aux deux.
Les effets génétiques de la selection
Les progrès actuels en génétique nous permettent de visualiser les effets de ces sélections sur le génome de la plante. On a ainsi pu identifier deux gènes responsables de la structure des plants de maïs, les gènes TB1 et TGA1. Ils sont respectivement responsables de l'architecture des ramifications latérales et de la mise en place de la glume.
La comparaison grâce au logiciel anagène de ces gènes nous permet de mieux comprendre ce sur quoi a porté la selection empirique.
Le gène TGA1
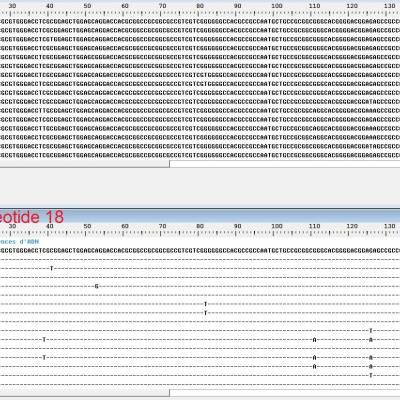
On observe une mutation commune à tous les maïs, une substitution au niveau du nucléotide 18 d'une cytosine par une guanine. Cette mutation entraîne la modification d'une lysine chez le téosinte par une asparagine chez le maïs dans la protéine codée par le gène TGA1. Cette modification entraîne une modification de la glume qui devient nettement plus petite.
Le gène TB1
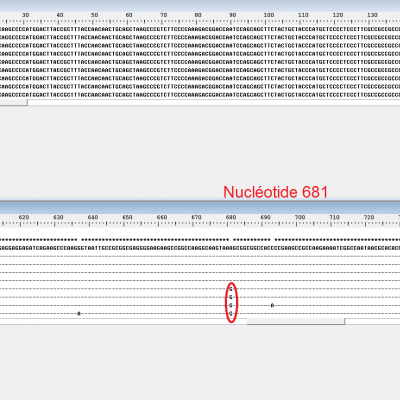
En réalisant une comparaison entre les gènes TB1 des maïs et des téosintes, on arrive bien à trouver une mutation commune à tous les maïs. Cependant, ceci est obtenu en réalisant un alignement avec discontinuité qui ne permet pas de dire que cette mutation est responsable d'une modification de l'activité de TB1. En revanche, celà permet de montrer que le gène TB1 est en réalité un gène homéotique dont les modifications de l'expression vont avoir une grande influence sur l'architecture de la plante. La surexpression de TB1 chez le maïs va entraîner l'inhibition de la croissance des ramifications latérales. On obtient une plante plus compacte.
Les expérience d'hybridation entre maïs "normal" et maïs mutant ont permis de montrer qu'un seul gène était responsable du port de la plante.
La génération F1 nous permet de dire que le phénotype normal est dominant (100% phénotype normal pour l'hétérozygotie).
Le croisement F1XF1 nous donne des résultats de type 75%/25% conforme à un brassage génétique avec un seul gène.
| Gamètes F1 | normal | mutant |
| normal |
normal/normal Phénotype normal |
normal/mutant Phénotype normal |
| mutant |
mutant/normal Phénotype normal |
mutant/mutant Phénotype mutant |
- Conclusion
Le maïs actuel est le fruit d'une longue selection des plants présentant le plus d'intérêts pour l'être humain. En se basant sur les caractéristiques les plus avantageuses pour sa nutrition, l'être humain a favorisé l'émergence d'une espèce domestiquée de la plante qui ne doit sa survie qu'aux apports nutritionnelle dont elle est capable. De ce fait, cette plante ne peut plus survivre dans un écosystème naturel mais ne peut s'épanouir qu'au sein d'un agrosystème sous la maîtrise de l'être humain. Les gènes impliqués dans ce processus ne sont pas forcément nombreux mais appartiennent généralement aux gènes architectes (homéotiques) Ainsi, ils peuvent modifier la plante de façon importante et durable.
![]() ?Tpmais (4.14 Mo)
?Tpmais (4.14 Mo)
![]() ?Tpmais (663.52 Ko)
?Tpmais (663.52 Ko)
- Le génie génétique au service de l'agriculture
- La génétique mendelienne
En 1856, gregor Mendel débute des expériences d'hybridation chez le pois. De ces expériences découlent des lois dites lois de Mendel qui permettent d'expliquer certaines transmissions génétiques de caractères d'une génération à une autre. On peut résumer ces lois:
Première loi : Loi d'uniformité des hybrides de première génération : aucune forme intermédiaire n'apparaît en F1 quand les parents sont de souches pures. Le concept de l'hérédité par mélange est réfuté.
Deuxième loi : Loi de pureté des gamètes : Les facteurs héréditaires se séparent dans les gamètes. Un gamète ne contient qu'un facteur de chaque caractère.
Troisième loi : Ségrégation indépendante des caractères héréditaires. Le cas pour les homozygotes.
Ces découvertes ont permis d'utiliser les hybridations pour "améliorer" les plantes cultivées, entendons par là sélectionner des variétés dont les caractères présentent un intérêt pour la nutrition de l'humanité (taille des fruits, goût, apport énergétique...) ou pour la facilité de culture ( résistance aux maladies ou aux ravageurs, faible encombrement...).
Nous pouvons illustrer l'application des travaux de Mendel par cet exemple :
Exemple de sélection après hybridation sur des plants de tomates
Dans une région au climat propice, on cultive deux variétés de tomates de lignée pure :
• l'une "A", à gros fruits
• l'autre "B", à petits fruits.
Les plants de la catégorie "A" se sont révélés sensibles à un champignon parasite : le Fusarium, qui
entraîne une baisse importante de production. En revanche, les plants de la variété "B" sont résistants à
ce champignon.
On demande à des agronomes de créer une nouvelle variété de plants de tomate donnant de gros fruits
et résistants au Fusarium. Ils réalisent une série de croisements entre les deux variétés de plants de
tomates "A" et "B".
A la première génération (F1), ils n'obtiennent que des plants de tomates résistants au Fusarium et qui
produisent des petits fruits.
Les chercheurs réalisent alors un autre croisement de la génération F1 avec des plants de la variété "A".
Ils obtiennent dans ces conditions à la deuxième génération (F2) les résultats suivants pour 1000 plants :
• 251 plants à petits fruits et résistants au Fusarium.
• 234 plants à petits fruits et sensibles au Fusarium.
• 270 plants à gros fruits et résistants au Fusarium.
• 245 plants à gros fruits et sensibles au Fusarium.
Peut on obtenir une lignée pure de plants de tomate donnant de grosses tomates résistantes? Si oui, préciser les étapes à suivre.
La question est plus complexe qu'il n'y parait. A première vue, il semble simple d'obtenir une lignée pure. Résolvons cet exercice de génétique dans les règles.
Les plants de tomate A donnent de gros fruits sensibles au champignon Fusarium. Nous allons noter ce phénotype [G;S]
Les plants de tomate B donnent de petits fruits résistants au Fusarium. Nous allons noter ce phénotype [P;R]
Les deux plants sont de lignée pure, on les considère donc homozygote.
En croisant ces pieds de tomate, on obtient une génération F1 hétérozygote qui présente un seul phénotype [P;R]. On en déduit que les phénotypes dominants sont P et R et que les phénotypes récéssif sont G et S. Nous allons donc nommer les gènes responsables des caractères:
Le gène de resistance au champignon sera nommé s. Il possède deux allèles : L'allèle s qui donne le phénotype S et l'allèle s+ qui donne le phénotype résistant.
Le gène de taille des fruits sera nommé g. Il possède deux allèles: L'allèle g qui donne le phénotype G et l'allèle g+ qui donne le phénotype P.
Donc, pour les plants A, on obtient: (gg;ss) ; [G;S]
Pour les plants B: (g+g+;s+s+) ; [P;R]
La génération F1 peut donc être caractérisée ainsi: (g+g;s+s) ; [P;R]
En effectuant un test-cross avec le parent double récessif (A), on obtient quatre phénotypes en F2 dont la proportion est d'environ 25% pour chaque phénotype, ce qui nous indique que les deux gènes sont indépendants.
|
g+;s+ |
g+s |
gs+ |
gs |
|
| g;s |
(g+g;s+s) [P;R] |
(g+g;ss) [P;S] |
(gg;s+s) [G;R] |
(gg;ss) [G;S] |
On obtient donc bien des plants résistants au Fusarium et donnant de gros fruits. Cependant, il ne s'agit pas d'une lignée pure mais d'une lignée hétérozygote.
On peut effectivement obtenir assez facilement une lignée pure en croisant les individus de F2 donnant de gros fruits et étant résistants. Cependant, seuls 25% des plants seront de lignée pure et il sera impossible de les distinguer des individus hétérozygotes. On peut donc obtenir des plants de lignée pure, mais pas 100% de plants de lignée pure.
L'application des découvertes en génétique a permis de passer d'une sélection empirique à une sélection scientifique.
- Les apports du génie génétique
Pour aller encore plus loin dans l'obtention de variétés présentant des caractères avantageux pour l'humanité, il est désormais possible d'insérer des gènes codant pour des caractères qui ne sont pas présents chez la plante. L'exemple suivant est encore pris chez le maïs.
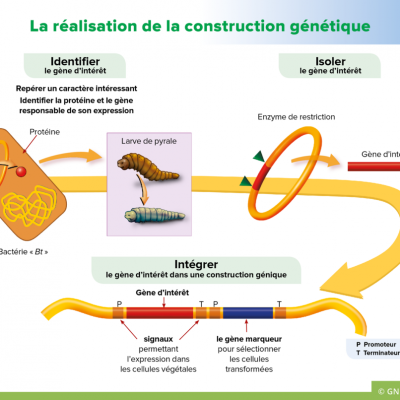
Le principal ravageur du maïs est une chenille, la pyrale (Ostrilia nubilalis). Afin de se débarrasser de cet insectes, on pulvérise de grandes quantité de pesticides sur les cultures. Il existe cependant un organisme produisant naturellement une toxine éliminant la pyrale : la bactérie Bacillus thuringiensis. Le gène permettant de coder la toxine est nommé Cry1Ab. Les chercheurs en génie génétique ont donc eu l'idée d'incorporer le gène Cry1Ab au patrimoine génétique du maïs.
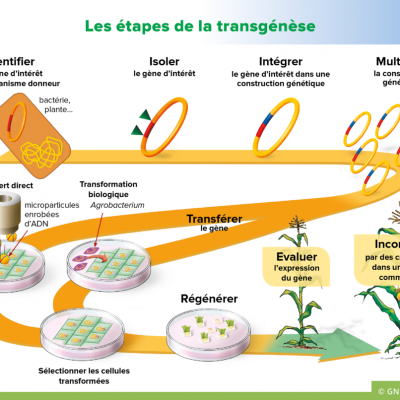
Les étapes de la transgénèse sont:
1. Repérer un caractère intéressant dans un organisme vivant et identifier la protéine.
2. Identifier et isoler le gène d'intérêt.
3. Réaliser et amplifier une construction génique
4. Transférer de l'ADN = Introduction d'un ADN étranger dans une cellule embryonnaire de Maïs.
5. Contrôler l'efficacité du transfert chez l'hôte.
6. Sélectionner des cellules exprimant le gène ajouté par tri.
Les outils de la transgénèse sont :
- Des enzymes de restriction capables de couper l'ADN en fonction de la séquence de nucléotide.
- Des plasmides bactériens, petites structures d'ADN circulaire possédant un facteur de transcription et capables de se reproduire rapidement. On connait précisément leur séquence.
- Une bactérie capable d'injecter les plasmides dans les cellules végétales : Agrobactérium tumefasciens
- Des gènes permettant la production de molécules de resistance aux antibiotiques et dont on connait précisément la séquence.
- Le gène d'intérêt Cry1Ab dont on connait précisément la séquence.
- Des antibiotiques
- Des cellules embryonnaires de maïs
A l'aide de ces outils, essayez de retrouvez les manipulations nécessaires à l'obtention d'un maïs génétiquement modifié pour produire naturellement la toxine contre la pyrale du maïs. Votre réponse peut prendre la forme d'un schéma, d'un texte ou d'un tableau.
- Première étape : Le caractère de résistance à la pyrale a été identifié chez la bactérie Bacillus thuringiensis. Il s'agit d'une molécule toxique pour la chenille.
- Deuxième étape : la molécule est codée par le gène Cry1Ab. Le séquençage nous permet de connaître la séquence du gène. On peut donc l'isoler en utilisant une enzyme de restriction qui va couper de part et d'autre du gène Cry1Ab. On se retrouve avec un gène isolé que l'on va pouvoir utiliser.
- Troisième étape : A l'aide de la même enzyme de restriction, on insère le gène Cry1Ab dans un plasmide bactérien en lui ajoutant des gènes de résistance aux antibiotiques. L'amplification des plasmides se fait le plus souvent par PCR, ce qui nous donne une grande quantité de plasmides porteurs d'un facteur de transcription, du gène de la toxine et de gènes de résistance aux antibiotiques.
- Quatrième étape : Le transfert de ce plasmide aux cellules de maïs va se faire en deux temps:
- Premier temps : on transforme les bactéries Agrobactérium tumefaciens avec les plasmides modifiés. Les bactéries qui ont bien incorporé le plasmide sont ensuite triée par culture sur milieu contenant des antibiotiques, ainsi, seules les bactéries disposant de gènes de résistance peuvent survivre et se multiplier (cinquième étape).
- Deuxième temps : On met en présence les cellules de maïs et les bactéries Agrobacterium tumefaciens afin qu'elles transfèrent le plasmide dans le maïs. On sélectionne ensuite les cellules de maïs génétiquement modifiée en les cultivant dans un milieu contenant des antibiotiques. Seuls les plants de maïs GM pourront se développer (sixième étape).
D'autres techniques de transgénèse existent, on peut ainsi bombarder les cellules à modifier par des microbilles (en or ou en tungstène) porteuses du gène d'intérêt.
Date de dernière mise à jour : 14/05/2020
Ajouter un commentaire