Chapitre 4: La commande réflexe des muscles
Introduction :
Les muscles sont les moteurs de nos mouvements. La contraction des muscles squeletiques permet d'induire un mouvement en tirant sur les tendons. Les articulations permettent ensuite le mouvement dans des directions préférentielles. Il existe ainsi tout un ensembles de muscles antagonites qui permettent le maintien de la posture, élément primordial du fait de notre bipédie. Cependant, les mouvements peuvent être le fruit de notre volonté via le système nerveux central ou résulter de commandes réflexe qui ne nécessite pas l'intervention de la conscience.
- Le réflexe myotatique
Problématique : Quels sont les mécanismes mis en œuvre lors d’un réflexe myotatique ?
La bipédie humaine exige un maintien permanent de l’équilibre. Pourtant, il n’est pas nécessaire de penser en permanence à garder cet équilibre pour rester debout. Si tel était le cas, il serait impossible de penser ou réfléchir tout en étant debout. Le maintien de l’équilibre est donc basé sur des phénomènes inconscients : les réflexes. Lors d'une visite de contrôle, le médecin peut vérifier le bon fonctionnement du système nerveux périphérique en appliquant un choc au niveau de certains tendons :
|
Nom du réflexe
|
Lieu du stimulus
|
Muscle stimulé
|
Articulation
|
Réponse induite
|
|
Rotulien
|
Tendon inférieur de la rotule
|
Quadriceps de la cuisse
|
Genou
|
Extension de la jambe
|
|
Achilléen
|
Tendon d'achille
|
Soléaire
|
Cheville
|
Extension du pied
|
|
Bicipital
|
Tendon au creux du coude
|
Biceps
|
Coude
|
Flexion de l'avant-bras
|
|
Tricipital
|
Tendon postérieur du coude |
Triceps
|
Coude
|
Extension de l'avant bras
|
Le principe est toujours le même : on réalise un stimulus (choc) au niveau d'un tendon impliqué dans une articulation puis on observe la réponse induite, flexion ou extension.
Nous allons essayer de comprendre le mécanisme du réflexe achilléen :
Observations cliniques :
| Données médicales | Observations/conséquences |
| Lésion de la partie supérieure de la moelle épinière (causée notamment par un grave traumatisme au niveau cervical) |
Paralysie Réflexe achilléen conservé |
| Lésion de la partie inférieure de la moelle épinière (causée notamment par un grave traumatisme au bas de la région dorsale) |
Paralysie Disparition du réflexe achilléen |
| Section accidentelle ou compression importante de la branche interne du nerf sciatique qui innerve les muscles du mollet | Disparition du réflexe achilléen |
| Dégénérescence de certaines structures sensorielles incluses dans le tissu musculaire (fuseau neuromusculaire) |
Absence de réflexe achilléen Maintien de la motricité volontaire |
| Dégénerescence des cellules musculaires (dans le cas des myopathies par exemple) | Absence de réflexe achilléen |
Identification des structures mises en jeu :
- Moelle épinière
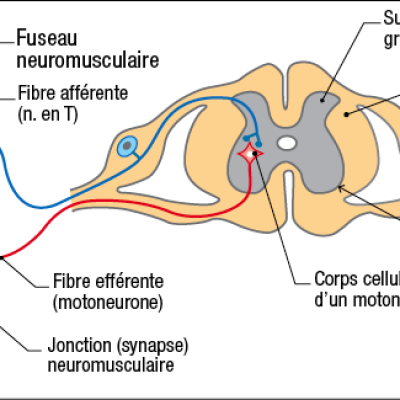
- Nerf sensoriel (fibre afférente) + fuseau neuromusculaire et nerf moteur (motoneurone) + plaque motrice (synapse neuromusculaire)
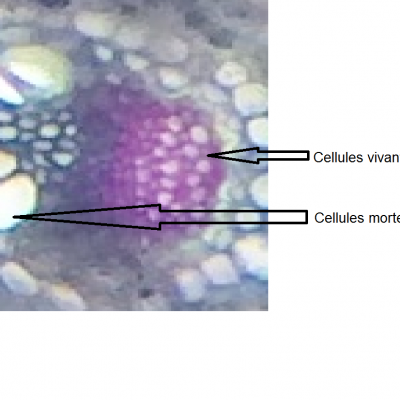
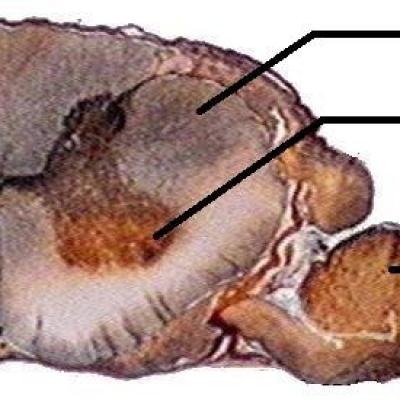
- Tendon d'Achilles ; Muscle soléaire (extenseur du pied) ; Muscle tibial antérieur (fléchisseur du pied)
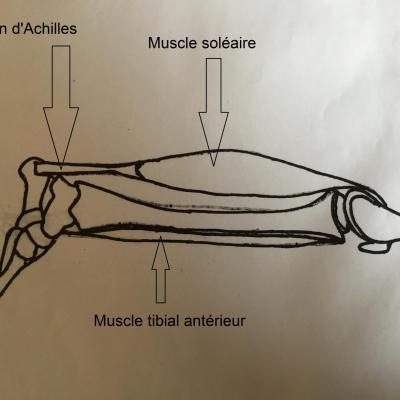
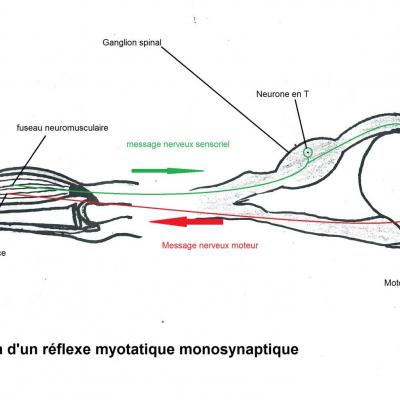
Le choc sur le tendon provoque une légère extension des fibres musculaires. Le fuseau neuromusculaire, constitué de fibres afférentes enroulées autour des fibres musculaires génèrent un message nerveux qui chemine jusqu'au corps cellulaire du neurone sensoriel (neurone en T) situé dans le ganglion spinal de la moelle épinière. Il est ensuite transmis au corps cellulaire du motoneurone positionné dans la substance grise. Le motoneurone génère alors un message nerveux qui va être conduit par l'axone (fibre efférente) de la corne ventrale jusqu'à la plaque motrice. Là, l'arrivée du message provoquera une contraction des fibres musculaires, le muscle va se raccourcir pour répondre à son élongation initiale. Sous l'effet de ce raccourcissement, le pied se tend. On parle de réflexe (le système nerveux central n'est pas impliqué comme le prouvent les expérience de sections) monosynaptique (il n'existe qu'une seule synapse entre le neurone en T et le motoneurone).
- Le message nerveux
La réponse réflexe repose sur la transmission d’un message nerveux depuis le récepteur sensoriel jusqu’à la moelle épinière et, en retour, de la moelle jusqu’au muscle. Ainsi, un signal de nature électrique se propage le long des fibres nerveuses, assurant une communication rapide entre organes. Ce message nerveux doit également être transmis d’un neurone à un autre ou d’un neurone vers un organe effecteur comme le muscle. Ces zones de connexion sont appelées synapses.
- Potentiels électriques et message nerveux.
Les neurones sont des cellules présentant un potentiel membranaire, également appelé potentiel de repos. Il repose sur l'existence d'un déséquilibre électrolytique entre l'intérieur de la cellule et l'extérieur de la cellule.
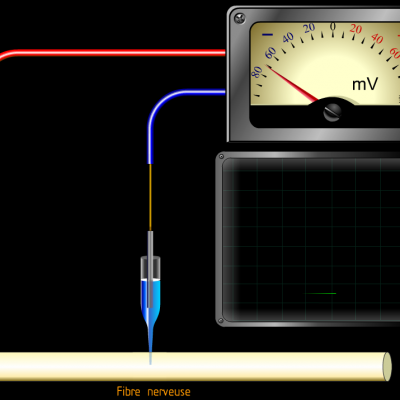
Entre l’électrode située à la surface de la fibre et l’électrode implantée dans la fibre, on observe une tension électrique négative d’environ -70mV. Ce qui signifie qu’il existe une accumulation de charges négatives à l’intérieur de la fibre.
L’expérience de Nernst (1864/1941) permet de déduire que les membranes sont semi-perméables aux ions. En effet, lorsque deux compartiments contenant des solutions de concentrations différentes sont séparés par une membrane perméable, des échanges d’ions se produisent visant à équilibrer les concentrations de part et d’autre de la membrane. On parle d’osmolarité. Les échanges se font donc toujours du compartiment où la concentration est la plus forte vers le compartiment où la concentration est la plus faible. On observe alors un transfert temporaire d’électron d’un compartiment à l’autre, un courant électrique. En réalité, la membrane est imperméable aux ions, mais il existe des canaux protéiques perméables aux ions potassium. Ils permettent une sortie des ions potassium sous l’effet du gradient de concentration. Ce déplacement associé à la présence de protéines chargées négativement à l’intérieur de la cellule permet d’obtenir une charge globalement négative à l’intérieur de la cellule, ce qui explique l’existence du potentiel de repos de -70 mV. Le déséquilibre doit être maintenu afin de permettre le maintien de cette polarisation. Il existe donc des pompes ioniques, nécessitant de l’énergie sous forme d’ATP, qui transportent les ions contre le gradient de concentration.

Ce déséquilibre entretenu est à l’origine de la rapidité des messages nerveux. Lors d’une stimulation, plusieurs phénomènes vont se succéder, permettant une variation instantanée du potentiel de membrane, appelée potentiel d’action.
- Les canaux calciques permettant le passage des ions sodium s’ouvrent et les ions Na+ entrent passivement dans la cellule sous l’effet du gradient de concentration. L’entrée massive de charges positives provoque une dépolarisation de la cellule, l’intérieur initialement négatif (-70 mV) devenant positif (+30 mV)
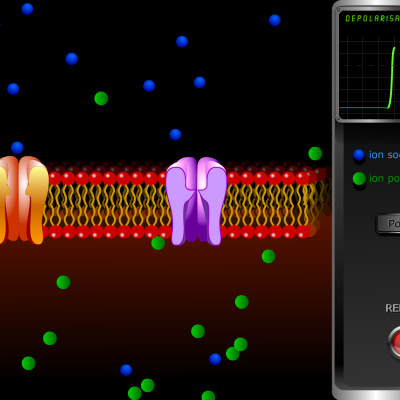
- Les canaux calciques se referment et les canaux potassiques s’ouvrent, permettant la sortie passive des ions K+ sous l’effet du gradient de concentration. La cellule se repolarise (-70 mV).
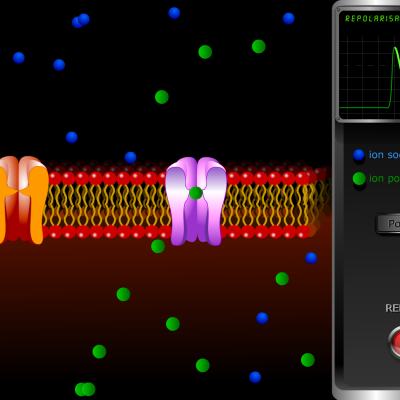
- L’ouverture des canaux potassique se poursuit et la cellule s’hyperpolarise, l’intérieur devenant plus négatif que ce qui est observé lors du potentiel de repos (-80 mV). Cette hyperpolarisation est une période réfractaire au cours de laquelle aucun nouveau potentiel d’action ne peut être engendré.
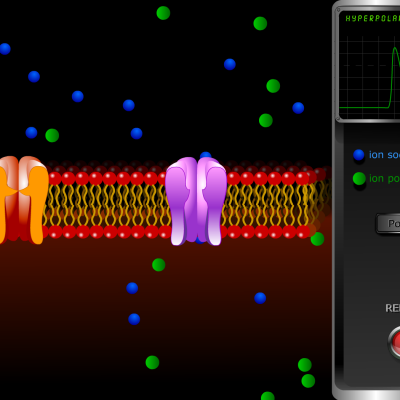
- Les canaux potassiques se ferment et la pompe ionique permet de rétablir le déséquilibre ionique qui engendre le potentiel de repos.
|
|
1 : Dépolarisation Entrée des ions Na+
2 : Repolarisation Sortie des ions K+
3 : Hyperpolarisation Sortie des ions K+
4 : Retour au potentiel de repos Pompes ioniques à ATP :entrée des ions K+, sortie des ions Na+ |
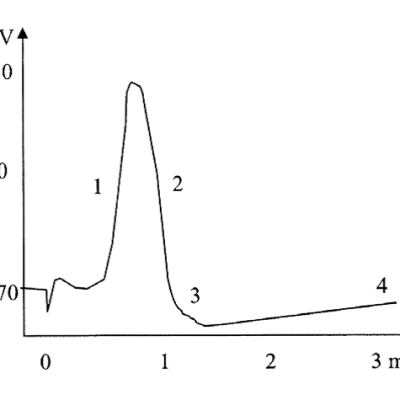
Les potentiels d’actions peuvent donc être vus comme des vagues d’électronégativité circulant à la surface des membranes des neurones. Ils ne sont créés que si une valeur seuil de déclenchement est atteinte, les potentiels d’action suivent donc la règle du tout ou rien :
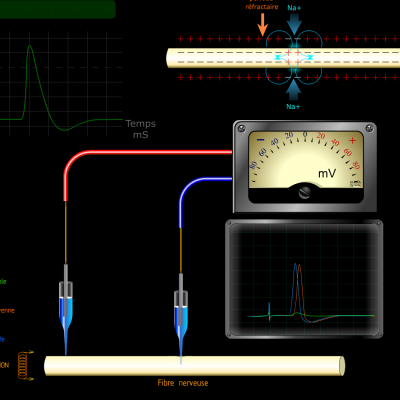
Si la valeur seuil n’est pas atteinte, aucun potentiel n’est engendré. Au-delà de la valeur seuil, un potentiel d’action d’intensité identique est créé.
Stimulus :
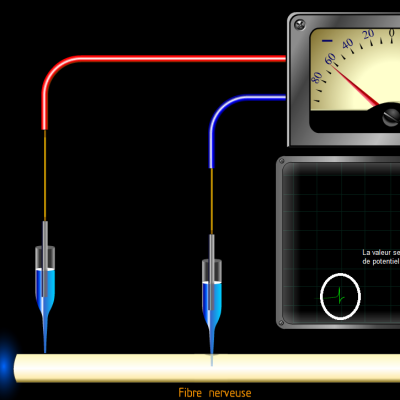
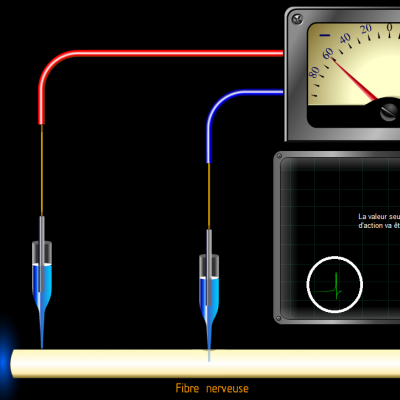
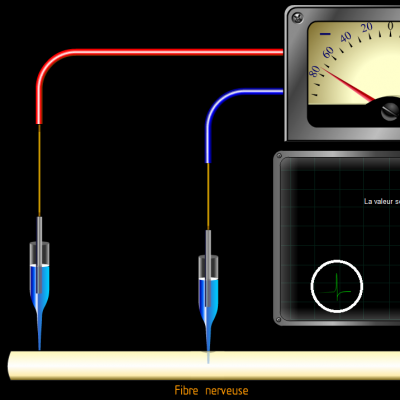
Potentiel d’action :
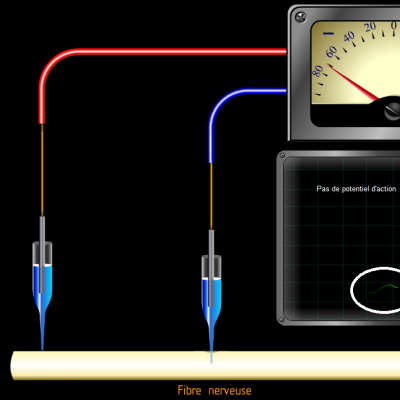
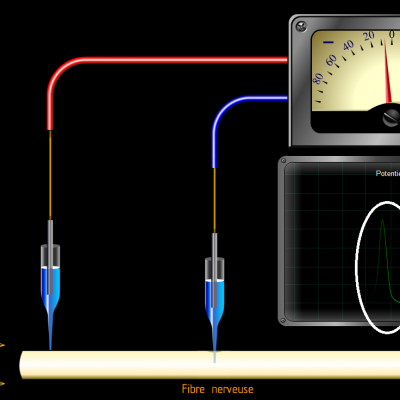
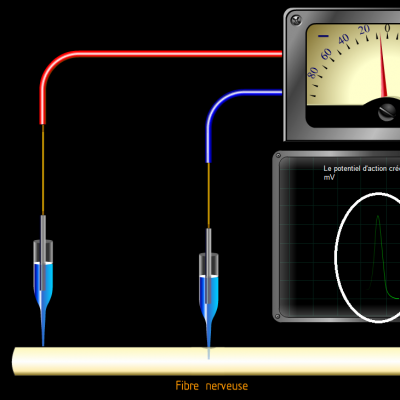
C’est donc la fréquence des salves de potentiels d’action qui reflète l’intensité du stimulus. Le message nerveux est donc codé par la fréquence des potentiels d’action induits par un stimulus. Plus le stimulus sera de forte intensité, plus la fréquence des potentiels d’action sera élevée.
- Les synapses
Les trajets que doivent parcourir les messages nerveux représentent de grandes distances pour des messages électriques d’aussi faible intensité. Afin de préserver le message nerveux, il existe donc des synapses entre neurones qui sont autant de relais permettant le maintien de l’intégrité du message nerveux. Au niveau de ces synapses, le message électrique est temporairement transformé en message chimique.
Nous allons nous appuyer sur l’exemple des synapses neuromusculaires pour étayer notre propos.
Les indiens d’Amazonie utilisent le curare, pour chasser leur proie en les paralysant avec des fléchettes projetées à l’aide de sarbacanes. Pour l’être humain, le curare est un poison qui provoque la mort par paralysie des muscles respiratoires. Au 19ème siècle, Claude Bernard étudia le mode d’action du curare et proposa qu’il empêche le contact entre les nerfs et le muscle. Le curare agirait donc au niveau de la synapse neuromusculaire sur le récepteur à l’acétylcholine.
Dans une synapse, l’arrivée d’un potentiel d’action déclenche la libération par l’élément présynaptique de molécules dans l’espace inter synaptique qui vont aller se fixer sur des récepteurs postsynaptiques. Ces molécules sont des neuromédiateurs (ou neurotransmetteurs).
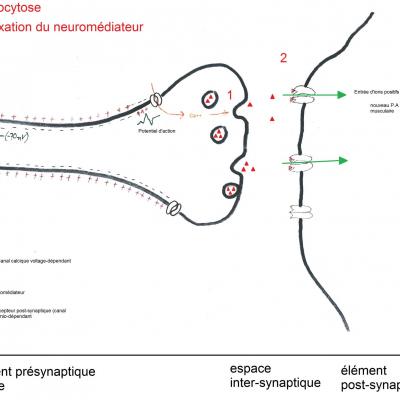
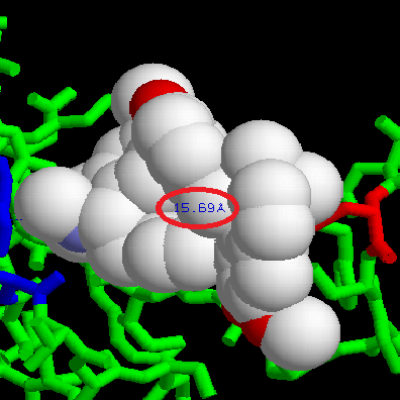
Dans le cas de la synapse neuromusculaire, le neuromédiateur est l’acétylcholine. Sa fixation sur un récepteur nicotinique provoque la contraction musculaire. La demi-vie d’un neuromédiateur est courte afin que les récepteurs puissent rapidement être à nouveau libres de déclencher une nouvelle réponse. Comme souvent en biologie, les synapses fonctionnent sur le mode clé/serrure. Or, certaines molécules présentent une structure tridimensionnelle proche des neuromédiateurs. Ainsi, la tubocurarine est une molécule capable de se fixer sur les récepteurs nicotiniques. Cependant, alors que l’acétylcholine se fixe temporairement aux récepteurs nicotiniques puis est rapidement dégradée, la tubocurarine reste fixée et empêche l’acétylcholine de venir se fixer sur les récepteurs. De plus, la fixation de la tubocurarine n’engendre aucune contraction musculaire. L’utilisation de Rastop nous permet de constater que lors de la fixation de l’acétylcholine, les acides aminés CYS 188 et TRP 145 sont séparés de moins de 10 Angstrom (10-10 nm). Le récepteur est activé et une contraction musculaire se produit.
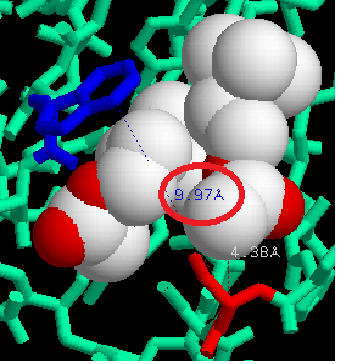
En revanche, la fixation de la tubocurarine entraîne une distance entre ces deux acides aminés de 15 angstrom. Le récepteur est bloqué mais pas activé, il ne peut donc y avoir de contraction musculaire. La tubocurarine est donc un antagoniste de l’acétylcholine.
![]() Tp message nerveux (713.66 Ko)
Tp message nerveux (713.66 Ko)
![]() Tp message nerveux (326.86 Ko)
Tp message nerveux (326.86 Ko)
Date de dernière mise à jour : 02/06/2020
Ajouter un commentaire