Chapitre 5
Chapitre 5 : Les relations entre organisation et mode de vie, résultat de l'évolution : l'exemple de la vie fixée chez les plantes
- Introduction et rappels
Schéma de l’organisation d’une plante
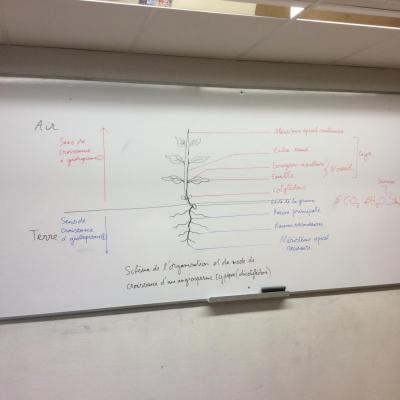
Les plantes sont des organismes autotrophes dont la particularité est de vivre fixée à l’interface entre deux millieux, le sol et l’atmosphère. De nombreuses adaptations se sont ainsi mises en place afin de pouvoir évoluer dans cet environnement particulier.
- Organisation de la plante
- Surfaces d’échange
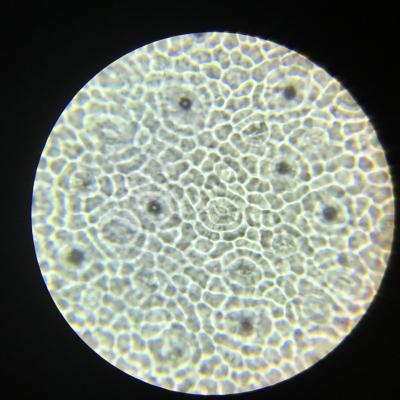
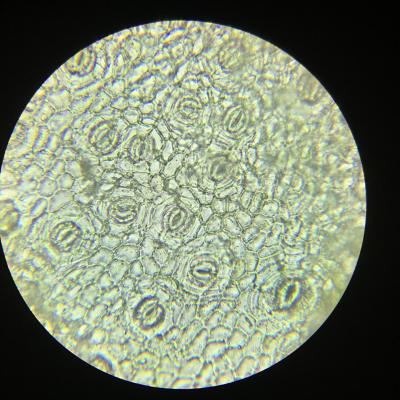
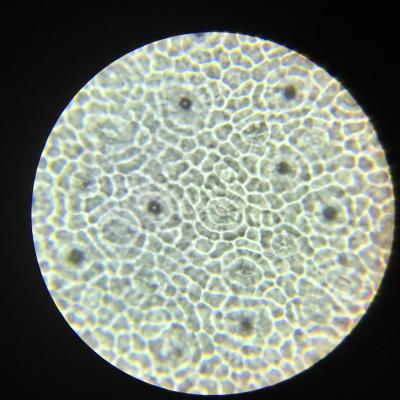
TP 1 Les surfaces d’échanges d’une plante avec l’atmosphère et avec le sol
![]() Tp sur les surfaces d echange (1.07 Mo)
Tp sur les surfaces d echange (1.07 Mo)
Résultats obtenus :
Bilan :
- Les plantes sont majoritairement des organismes autotrophes, capables de produire leur propre matière organique à partir de matières minérales, eau et dioxyde de carbone et d’énergie lumineuse. En effet, la photosynthèse peut être modélisée sous forme d’une équation bilan : 6CO2 + 6H2O à C6H12O6 + 6O2.
- La position de la plante à l’interface des deux milieux que sont l’atmosphère et le sol lui permet de profiter des ressources disponibles dans les deux milieux : dioxyde de carbone dans l’atmosphère et eau dans le sol. Afin d’optimiser les échanges, la forme des organe et deux structures spécialisées se sont mises en place.
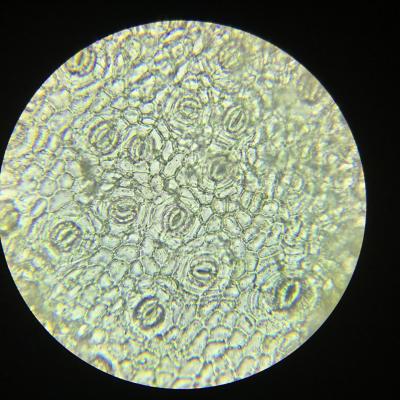
- Les feuilles sont plates et fines afin de pouvoir capter au mieux les rayons solaires et disposent de nombreux stomates dont le rôle est de capter le dioxyde de carbone. Le calcul de leur densité permet d’obtenir une estimation des capacités d’échanges gazeux de la feuille.
- Les racines sont longues, d’un diamètre relativement faible et cylindriques afin d’optimiser leur contact avec le sol dans le but de capter l’eau. La présence de nombreux poils absorbants démultiplie encore la capacité de la plante à absorber eau et sels minéraux du sol.
L’eau est donc captée dans le sol par les racines, le dioxyde de carbone par les stomates au niveau des feuilles. On peut donc désormais se demander comment ces deux substances peuvent se rejoindre au niveau de la zone où a lieu la photosynthèse, le parenchyme foliaire.
- Les systèmes conducteurs dans la plante
L’eau et les sels minéraux proviennent du sol où les poils absorbants présents dans les racines les absorbent. Ces substances doivent donc rejoindre les feuilles où s’effectuent la captation du dioxyde de carbone et la photosynthèse. Les matières organiques et les métabolites produits doivent alors être distribués dans l’ensemble de la plante. On peut donc penser qu’il existe un double système de circulation au sein de la plante, un système ascendant, capable de conduire la sève brute (eau + sels minéraux) des racines aux feuilles et un système descendant qui va amener la sève élaborée (eau + substances photosynthétiques) aux parties inférieures de la plante.
TP sur les systèmes conducteurs de sève
![]() Tp sur les systemes conducteurs de seve 1 (17.87 Ko)
Tp sur les systemes conducteurs de seve 1 (17.87 Ko)
Objectifs notionnels de la séance :
- Observer la présence de vaisseaux conducteurs de la sève brute de la racine à la feuille
- Comprendre et observer l’existence de vaisseaux conducteurs de la sève élaborée
- Comprendre et observer la répartition des vaisseaux au sein de la plante
Objectifs de capacités expérimentales :
- Réaliser des coupes fines de végétaux
- Réaliser avec soin et observer une lame microscopique
Première manipulation : repérage longitudinal des vaisseaux conducteurs de sève brute
Matériel :
Tige de céleri branche placées depuis plusieurs jours dans de l’eau colorée au bleu de méthylène, lames de rasoir, pince fine, lame, lamelles.
Manipulation :
- Couper transversalement un fragment de tige de longueur 1 cm. Le placer sur une lame.
- À l’aide d’un scalpel, inciser ce fragment dans sa longueur et l’ouvrir.
- À l’aide d’une pince fine, isoler dans sa longueur une des structures contenant l’eau colorée.
- Placer cette structure entre lame et lamelle dans une goutte d’eau.
- Observer au microscope et repérer les vaisseaux.
Deuxième manipulation : observation des vaisseaux en coupe transversale.
Matériel :
Tiges ou pétioles de lierre
Pince fine, lame de rasoir
Lames, lamelles, verres de montre
Eau de javel, acide acétique 10%, Carmino-vert de Mirande.
Manipulation :
- Prélever la tige et la placer sur une lame.
- Réaliser une coupe transversale de la tige (fendre dans le sens de la largeur) à l’aide d’une lame de rasoir.
- Déposer les coupes de tiges dans un verre de montre et recouvrir d’eau de javel. Laisser agir 20 min.
- Retirer les tiges avec la pince fine et les rincer dans un 2ème verre de montre contenant de l’eau.
- Placer les tiges dans un 3ème verre de montre contenant de l’acide acétique (1 à 2 min).
- Rincer à nouveau
- Placer les tiges 30 sec dans un verre de montre contenant du carmin-vert d’iode, puis rincer.
- Placer la (ou les) section(s) de tige entre lame et lamelle dans une goutte d’eau.
- Observer au microscope et repérer les vaisseaux.
Troisième manipulation : observation de coupe longitudinale de feuille et repérage des vaisseaux.
Matériel :
Feuille de poireau
Eau de javel à 2% dans l’eau distillée, Acide acétique à 1% dans l’eau distillée (Eau acétique), Carmino vert de mirande.
Verres de montre pour la double coloration
Pinces fines
Lames et lamelles
Manipulation :
1. Couper des morceaux de 4 à 5 cm dans une feuille de poireau bien verte.
2. Mettre ces morceaux à bouillir dans de l'eau pendant 10 minutes.
La cuisson permet de séparer facilement les tissus de la feuille.
3. À l'aide d'une pince fine dégager, en tirant, les tissus situés dans les nervures.
4. Placer 2 à 3 échantillons dans de l'eau de javel pendant 20 minutes.
5. Rincer dans de l'eau pendant quelques secondes
6. Placer l'échantillon dans de l'eau acétique pendant 3 à 4 minutes
7. Placer l'échantillon dans le colorant carmin vert d'iode pendant 3 minutes
8. Bien rincer l'échantillon dans de l'eau
9. Placer l'échantillon bien rincé dans une goutte d’eau, couvrir l'échantillon d'une lamelle
10. Observer au microscope et repérer les vaisseaux.
Remarque :
La coloration au carmino-vert de Mirande n'est pas une coloration spécifique de la lignine et de la cellulose : ces colorants ont seulement une affinité plus prononcée vis à vis de ces composés chimiques. Ainsi, en présence du seul carmin aluné tout est coloré en rouge, et réciproquement tout est coloré en vert par le vert d'iode : c'est l'association des deux colorants qui aboutit à une coloration différentielle des tissus (le xylème est coloré en vert).
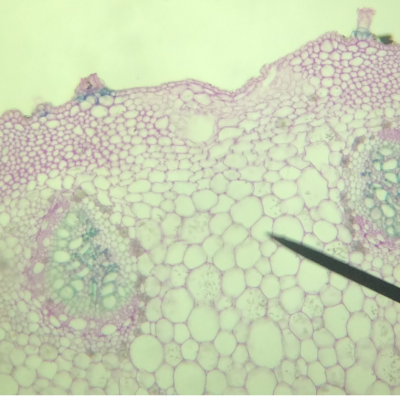
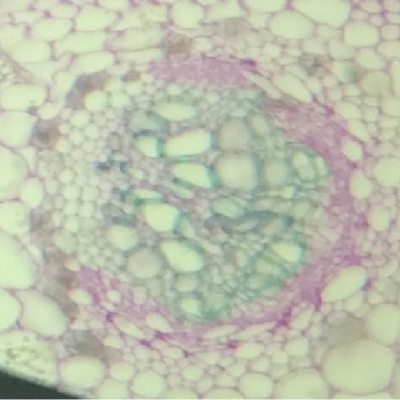
Résultats obtenus :
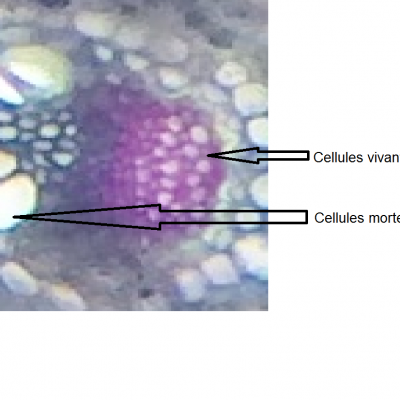
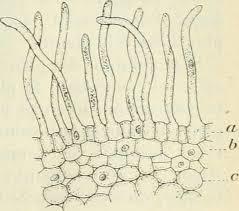
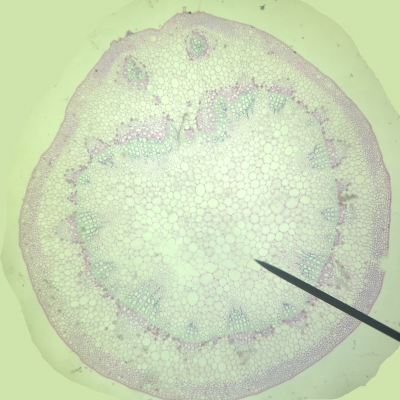
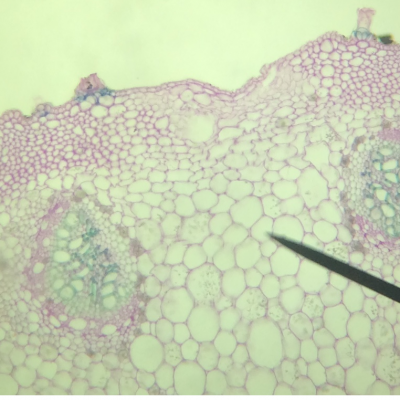
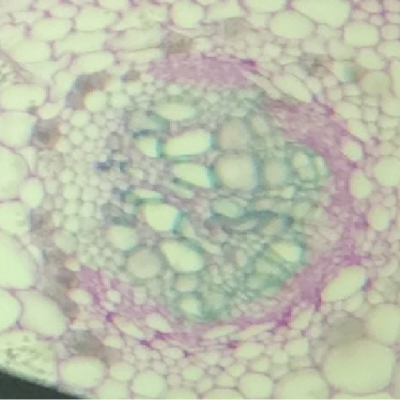
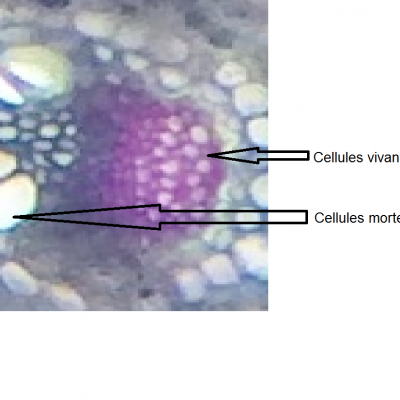
- On observe deux types de vaisseaux conducteurs :
Des vaisseaux qui semblent vides à l’intérieur et qui présentent des parois très épaisses colorés en bleu-vert.
Des vaisseaux plus fins, colorés en rose, constitués de cellules vivantes.
- Les vaisseaux vides sont ceux dans lesquels l’eau colorée a réussi à monter par capillarité.
|
|
|
|
|
|
Interprétations :
Les vaisseaux colorés en vert sont des cellules vides et mortes qui permettent une circulation de la sève brute (l’eau et les sels minéraux) sous pression ou sous tension grâce à leurs renforts en lignine dans les parois. La circulation peut donc y être ascendante.
Les vaisseaux colorés en rose sont des cellules vivantes qui comportent de la cellulose. Elles permettent la circulation de la sève élaborée (riche en matières organiques) par gravité, des feuilles vers les racines, donc majoritairement descendante.
Bilan :
Les plantes disposent d’un double système de circulation :
- Les vaisseaux du xylème sont constitués de cellules mortes renforcées grâce à leurs parois lignifiées. Elles peuvent donc supporter la pression nécessaire à la montée de la sève brute pendant la nuit et la tension qui accomplit la même chose pendant la journée.
- L’eau et les sels minéraux arrivent ainsi dans les feuilles où s’effectue la photosynthèse. Ce métabolisme permet de réduire le dioxyde de carbone atmosphérique dans des molécules organiques comme le glucose par exemple par l’intermédiaire de l’énergie lumineuse.
- Les métabolites produits dans les organes aériens sont ensuite acheminés vers les parties basses de la plante par l’intermédiaire des vaisseaux du phloème qui sont constitués de cellules vivantes, non lignifiées.
- Protection contre les agressions du milieu
De par leur mode de vie fixée, les plantes ont dû développer des moyens de protection originaux contre les agressions de leurs milieux de vie. Dans un premier temps, nous allons nous intéresser à la protection contre la sécheresse.
TP sur les systèmes de protection contre les agressions du milieu
Les plantes étant des organismes fixées, elles ne peuvent se déplacer pour pouvoir satisfaire leurs besoins en eau. Si cela ne semble pas être un problème dans les milieux humides, cela peut s’avérer mortel si la plante doit faire face à un milieu sec (déserts) ou si l’eau n’est pas aisément accessible (ruissellement important, milieu venté et ne retenant pas l’eau).
|
L’Oyat des dunes (Ammophilia arenaria) se développe, comme son nom l’indique, sur un sol très sableux, inhospitalier et incapable de retenir l’eau de pluie. De plus, le climat souvent venteux tend à dessécher les plantes. |
|

|
La Giroflée des dunes ou Matthiole à feuilles sinuées (Matthiola sinuata) est une plante herbacée bisannuelle de la famille des Brassicacées poussant sur les sables des côtes méditerranéenne et atlantique. |
|

|
Le muguet (Convallaria majalis) est lui une plante des sous-bois, il ne s’installe jamais sur des milieux secs. |
|

Proposer une stratégie de résolution permettant de répondre au questionnement :
Montrez que l’anatomie des feuilles permet de limiter les pertes d’eau, assurant ainsi une bonne adaptation aux milieux de vie.
Objectifs notionnels de la séance :
- Observer la présence de structures spécialisées dans les feuilles.
- Comprendre le fonctionnement et l’intérêt de ces structures dans le cadre d’une adaptation à un milieu où l’approvisionnement en eau est difficile.
- Constater que les stratégies peuvent être différentes malgré un objectif similaire : réduire les pertes d’eau pour la plante.
Objectifs de capacités expérimentales :
- Réaliser des coupes fines de végétaux
- Réaliser avec soin et observer une lame microscopique
Matériel :
Feuilles de giroflée des dunes, feuille d’oyat
Préparation microscopique de feuilles de muguet
Lames de rasoir, pince fine.
Lames, lamelles, verres de montre.
Eau de javel, acide acétique 10%, Carmino-vert de Mirande.
Protocole :
Manipulation :
- Prélever la zone médiane d’une feuille et la placer entre deux lames.
- Réaliser une coupe transversale de la feuille (fendre dans le sens de la largeur) en passant par le centre à l’aide d’une lame de rasoir.
- Déposer les coupes de feuilles dans un verre de montre et recouvrir d’eau de javel. Laisser agir 5 min.
- Retirer les coupes avec la pince fine et les rincer dans un 2ème verre de montre contenant de l’eau.
- Placer les coupes dans un 3ème verre de montre contenant de l’acide acétique (1 à 2 min).
- Rincer à nouveau
- Placer les coupes 30 sec dans un verre de montre contenant du carmin-vert d’iode, puis rincer.
- Placer la (ou les) section(s) de feuilles entre lame et lamelle dans une goutte d’eau.
- Observer au microscope et repérer les vaisseaux.
Remarque :
La coloration au carmino-vert de Mirande n'est pas une coloration spécifique de la lignine et de la cellulose : ces colorants ont seulement une affinité plus prononcée vis à vis de ces composés chimiques. Ainsi, en présence du seul carmin aluné tout est coloré en rouge, et réciproquement tout est coloré en vert par le vert d'iode : c'est l'association des deux colorants qui aboutit à une coloration différentielle des tissus (le xylème est coloré en vert).
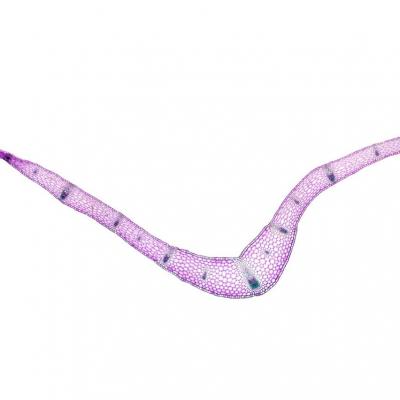
Résultats obtenus :
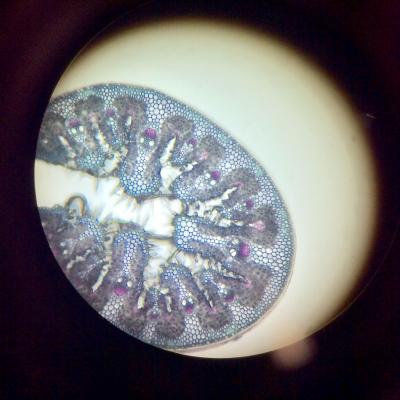
Oyat
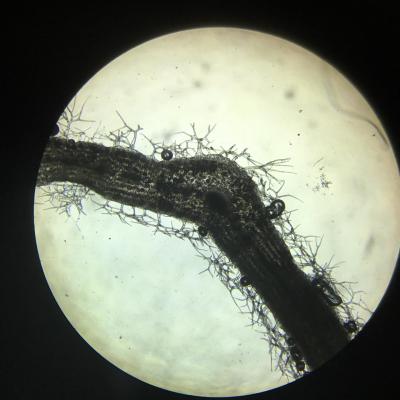
Giroflée des dunes
Muguet
Les TP à télécharger
![]() Tp sur les systemes de protection secheresse2 (210.29 Ko)
Tp sur les systemes de protection secheresse2 (210.29 Ko)![]() Tp sur les systemes de protection secheresse2 (151.48 Ko)
Tp sur les systemes de protection secheresse2 (151.48 Ko)
Observations:
Oyat:
- On observe une feuille enroulée sur elle-même de manière à ce que la face interne soit protégée.
- La face externe est recouverte d’une pellicule imperméable et présente des cellules dont les parois sont renforcées avec de la lignine.
- La face interne présente des zones de cellules dont les parois ne sont pas renforcée et qui sont constituées de cellulose.
- On observe également des cellules pilifères.
Giroflée du désert :
- La feuille présente des grandes cellules dans le parenchyme interne.
- Les deux faces comportent des cellules pilifères.
- Les poils sont ramifiés.
- Malgré les précautions et le soin apporté à la coupe, il est quasiment impossible de ne pas emprisonner de bulles d’air lors du processus d’obtention de l’échantillon.
- La feuille est aplatie mais de petite taille.
Muguet :
- La feuille est constituée de cellules dont les parois sont constituées de cellulose, pas d'adaptations particulières. (= témoin).
- La feuille est aplatie et présente une vaste surface d'échange.
Interprétations:
Les feuilles d’oyat présentent la particularité de pouvoir s’enrouler de manière à ne présenter que la face inférieure. La face supérieure est ainsi maintenue à l’abri des courants d’air qui pourraient dessécher la plante. Le mécanisme d’enroulement est dû à la présence de cellules rigides incapable de modifier leur volume sur la face inférieure. En revanche, certaines cellules de la face supérieure ne disposent pas de renforts de lignine et peuvent se gorger d’eau. Ce faisant, leur volume augmente et la feuille se déroule. Les poils présents à la surface permettent de conserver prisonnière l’eau accumulée pendant les périodes où l’atmosphère est humide.
Les feuilles de giroflée des dunes sont constituées de grandes cellules qui peuvent emmagasiner de l’eau. Les nombreux poils ramifiés jouent un double rôle. D’une part, ils jouent le rôle de réserve d’eau supplémentaire lorsque l’atmosphère est humide. D’autre part, ils peuvent également jouer le rôle d’isolant et éviter un trop grand dessèchement de la plante lorsque l’atmosphère est très sèche.
Conclusion :
Les plantes possèdent des moyens de lutter contre le dessèchement. Elles peuvent garder l’eau en réserve dans des cellules ou via des poils, elles peuvent modifier leur forme pour éviter des pertes trop importante d’eau, on sait aussi que certaines plantes ont des racines qui peuvent servir de réserves.
TD : défense contre les prédateurs.
![]() Acaciagirafes (781.93 Ko)
Acaciagirafes (781.93 Ko) ![]() Acaciagirafes (621.92 Ko)
Acaciagirafes (621.92 Ko)
![]() Acacia corrige (15.79 Ko)
Acacia corrige (15.79 Ko) ![]() Acacia corrige (122.82 Ko)
Acacia corrige (122.82 Ko)
Bilan :
Les plantes vivant fixées, elles n’ont pas la possibilité de se déplacer pour trouver des conditions favorables en cas de modification de leur milieu de vie ou d’attaques de prédateur. Des adaptations se sont développées afin de lutter contre les diverses agressions dont elles peuvent être la cible. Leurs organes peuvent présenter des modifications structurelles afin de répondre aux défis que sont la dessiccation, le feu, l’excès d’eau, les éléments chimiques et les prédateurs.
On peut classer ces adaptations en trois grandes catégories :
- Mécanique : Enroulement des feuilles (oyat), présence d’épines (rosier), de poils urticants (orties), mécanismes de torsions ou de fermeture rapide (tournesol, dionée)
- Chimique : Présence de substances toxiques (cigüe, aconit), diffusion de gaz (acacia), développement de substances amères (tanins, et alcaloïdes)
- Biologique : développement de symbioses (soja, trèfle, acacia)
- Reproduction chez les angiospermes
La vie fixée représente également une gageure dans le cadre de la reproduction. Chez les plantes appartenant au clade des angiospermes, la solution évolutive a été le développement de la fleur.
- Organisation de la fleur chez les angiospermes
TP sur l’organisation d’une fleur d’angiosperme
Proposer une stratégie de résolution permettant de répondre au questionnement :
Montrez que l’anatomie des fleurs est organisée en couronnes concentriques et déterminez sa mise en place génétique.
Objectifs notionnels de la séance :
- Observer la présence de 4 verticilles concentriques.
- Comprendre la mise en place génétique de cette structure.
Objectifs de capacités expérimentales :
- Réaliser une dissection florale
- Réaliser avec soin et observer une lame microscopique
Matériel :
- Fleur d’angiosperme
- Pince fine.
- Loupe
- Lames, lamelles, verres de montre.
- Scotch, feuille blanche
Protocole :
- Dissection florale
Manipulation :

- Détermination de la morphogénèse florale.
La mise en place de la fleur est déterminée génétiquement. On a pu identifier trois classes de gènes homéotiques, A, B et C qui interviennent dans les cellules qui composeront la fleur au niveau des bourgeons. Si ces trois classes de gènes sont inactivées, alors la fleur est composée de simples feuilles.
On a pu expérimentalement inactiver ces gènes de façon séquentielle.
Voici les résultats obtenus :
|
|
|
|
|
|
|
|
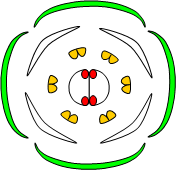
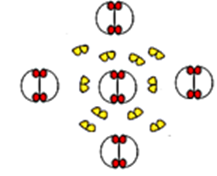
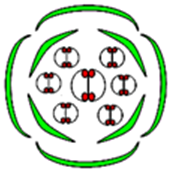
Enfin, on a pu déterminer que si les gènes de classe A sont inactivés, les gènes de classe C s’expriment à leur place et inversement. Cette exclusion réciproque ne concerne pas les gènes de classe B.
![]() Tp organisation florale (338.39 Ko)
Tp organisation florale (338.39 Ko) ![]() Tp organisation florale (371.71 Ko)
Tp organisation florale (371.71 Ko)
Résultats obtenus:
J'ai décidé de travailler sur une fleur de poirier, dites moi sur quelles fleurs vous avez travaillé si vous voulez une correction personnalisée.

Après prélèvement d'une fleur de poirier, on procède à sa dissection de l'extérieur vers l'intérieur.
Les premières pièces que l'on enlève sont les sépales qui constituent le calice. Dans la fleur de poirier, on en compte 5, de petite taille et de couleur brun-vert. Leur aspect est un peu pelucheux du fait de la présence de poils qui servaient de protection au bourgeon floral.
Nous trouvons ensuite les pétales, au nombre de 5 également et qui constituent la corolle. Ils sont de couleur blanche, lisse et plus grands que les sépales.
L'ensemble de ces deux premiers cercles est appelé périanthe. Ces pièces sont stériles et ont pour rôle de protéger les pièces fertiles.
Le cercle suivant est appelé androcée et est constitué de 20 étamines.Cest étamines sont constituées d'un filet et de deux sacs, les anthères, qui portent les grains de pollen.
Enfin, au centre de la fleur, le gynécée qui se compose d'un ovaire surmonté de 5 styles. La coupe de cet ovaire révèle 5 loges, les carpelles, dans lesquels on trouve les ovules.
On peut donc, après cette dissection, établir le diagramme floral ainsi que la formule florale.
Photographie de la dissection:

Détail de l'ovaire

Formule florale : 5S+5P+20E+5C
Mise en place génétique:
Les gènes homéotiques au niveau de l'ébauche florale fonctionnent ainsi:
Le premier verticille, celui des sépales ne voit s'exprimer que le gène A
Le deuxième verticille, celui des pétales, subit l'action des gènes Aet B
Le troisième verticille est caractérisé par l'action des gènes B et C ce qui permet l'apparition des étamines.
Le quatrième verticille, celui des carpelles est formé par l'action unique du gène C.
Bilan :
Chez les angiospermes, les fleurs sont organisées en quatre couronnes concentriques, les verticilles, qui se succèdent de la périphérie vers le centre toujours dans le même ordre. Les deux couronnes externes forment le périanthe et protègent les deux couronnes internes qui contiennent les organes reproducteurs :
Les sépales, le plus souvent verts et ayant l'apparence de petites feuilles, mais parfois colorés, constituent le calice.
Les pétales de formes et de couleurs très diverses composent la corolle.
Les étamines, organes mâles de la fleur, sont faites d'une fine tige, le filet, portant des sacs à pollen, les anthères.
Le pistil, organe femelle de la fleur, contient des ovules répartis dans plusieurs loges nommées carpelles.
L'observation de mutants montre parfois des modifications des couronnes de pièces florales. On a ainsi pu déterminer l'existence de gènes du développement floral. Il en existe 3 classes (A, B et C) qui s'expriment spécifiquement selon des territoires et déterminent le développement des ébauches de pièce florale en sépale, pétale, étamine et carpelle.
- Rencontre des gamètes
La fleur est donc organisée afin de faciliter la reproduction. Les fleurs d'angiospermes sont souvent hermaphrodites mais il existe de nombreux mécanismes qui empêchent l'autofécondation. La fécondation croisée est donc favorisée, évitant un apauvrissement trop important de la diversité génétique. Cependant, cette fécondation croisée impose que les grains de pollen puissent être transportés.
On cherche donc à mettre en évidence l'existence de structures ou de caractéristiques au niveau des grains de pollen ou de la fleur qui peuvent nous renseigner sur le mode de transport des grains de pollen.
Proposer une stratégie de résolution permettant de répondre à ce questionnement.
Matériel : fleur de colza
Objectifs notionnels de la séance :
- Observer les structures présentes sur le grain de pollen.
- Observer les caractéristiques de la fleur permettant la pollinisation.
Objectifs de capacités expérimentales :
- Réaliser une dissection florale
- Réaliser avec soin et observer une lame microscopique
Ressources :
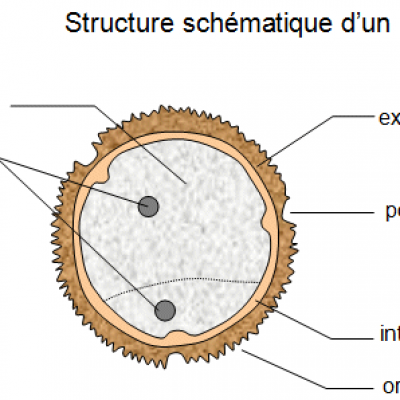
Document 1 : Structure schématique d'un grain de pollen.
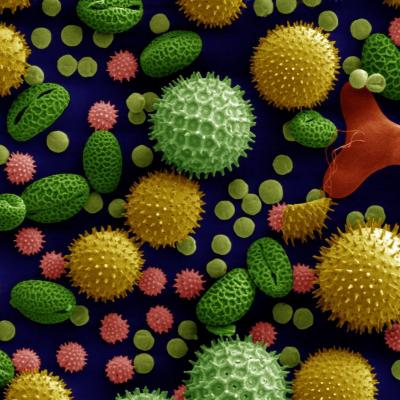
Document 2 : Quelques grains de pollen de 4 angiospermes.
Ressources complémentaires :
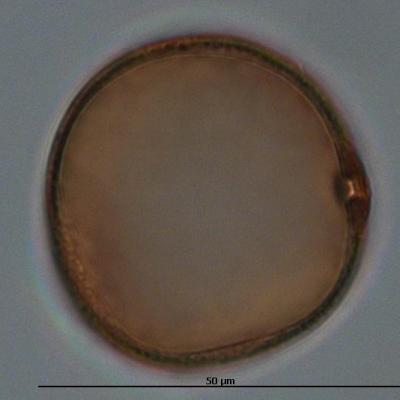
Pollen de poacée (transport par le vent)
Pollen de camomille (transport par des insectes)
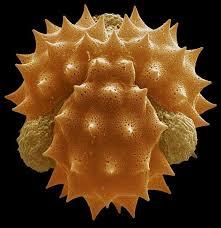
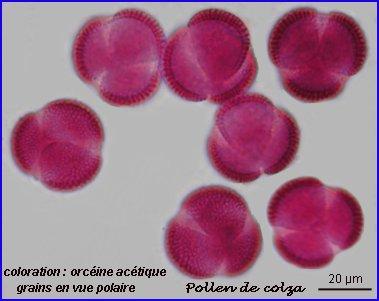
Résultats obtenus:
Dissection de la fleur de colza

Pollen de la fleur de colza
Et les TP version word et PDF
![]() Tp pollen v2 (560 Ko) ;
Tp pollen v2 (560 Ko) ; ![]() Tp pollen v2 (501.23 Ko)
Tp pollen v2 (501.23 Ko)
Observations :
La fleur de colza est une petite fleur jaune odorante. (Observée aves des rayons UV, on observe une coloration différente qui mène aux nectaires.) Ces nectaires sont au nombre de 4 et sont situés à la base de la corolle, près du pistil.
Les grains de pollen présente une forte ornementation et sont de taille moyenne.
Interprétations :
La fleur de colza est organisée afin de pouvoir être pollinisée par les insectes (entomogamie) La présence des nectaires à la base de la fleur oblige les pollinisateurs à se trouver en contact avec les étamines. L'ornementation des grains de pollen leur permet d'être captés par les poils des pollinisateurs, ce qui est facilité par leur taille moyenne. (Les abeilles, principaux pollinisateurs, sont en outre attirés par la couleur et l'odeur de la fleur et guidés vers les nectaires par la coloration que l'on peut observer en observant la fleur sous les UV.)
Bilan
Le transport des grains de pollen est rendu indispensable par l'exigence de fécondation croisée qui seule permet de maintenir la diversité génétique. Il existe plusieurs medias pour ce transport
Le vent (anémogamie) chez le chêne, le chataignier, le bouleau. Les grains de pollen sont alors plutôt de petite taille, avec une faible ornementation et parfois des ballonets aérifères. Cette stratégie nécessite une production massive de pollen.
L'eau (hydrogamie) chez la zostère. Dans ce cas, le grain de pollen est très allongé.
La zoogamie enfin chez la majorité des espèces (90%). Ce type de transport est beaucoup plus spécifique. Les fleurs sont alors attractives pour les pollinisateurs. Elles sont colorées, odorantes et produisent des substances nutritives.
La zoogamie exige donc une coévolution entre la plante d'une part et son pollinisateur d'autre part. L'association peut être si étroite que certaines plantes ne peuvent être pollinisées que par une seule espèce de pollinisateur (vanille et Euglossa viridissima ; figuier et blastophage).
- Formation et dispersion des graines
![]() Tddisp (3.62 Mo)
Tddisp (3.62 Mo)
![]() Tddisp (1.06 Mo)
Tddisp (1.06 Mo)
Une fois fécondée par le pollen, le pistil se transforme en fruit qui contient les graines. Ces graines doivent ensuite être transportées afin d'assurer la dissémination de l'espèce. Comme pour la pollinisation, les plantes disposent de plusieurs moyens de dispersion des graines afin d'assurer la colonisation d'autres milieux de vie. Nous avons vu que les grains de pollen disposent de structures spécifiques qui permettent leur transport par l'air, l'eau ou d'autres êtres vivants. En est-il de même pour le transport des graines?
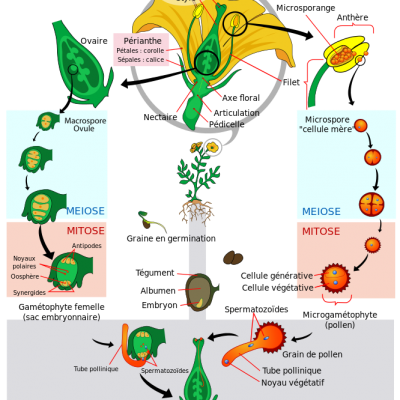
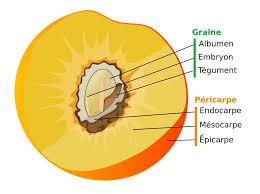
En étudiant le cycle de vie d'une plante, on observe que lors de la reproduction, le pollen se dépose sur le stigmate de la fleur et germe pour aller féconder les ovules contenus dans le pistil. Après la pollinisation, la fleur se transforme. La plupart des pièces florales fanent (sépales, pétales et étamines). La paroi de l’ovaire devient la paroi du fruit nommée péricarpe qui est divisé en trois parties : l’épicarpe (la peau de la prune), le mésocarpe (la chair) et l’endocarpe (le noyau). Les ovules fécondés deviennent les graines comprenant l’embryon et des cotylédons (feuilles chargées de réserves organiques (l’embryon de prune possède deux cotylédons, le prunier est donc une dicotylédone). Il existe de nombreuses formes de fruits.
En absence de transport, les grainent germent au pied de la plante mère et se retrouvent donc privées de nombreuses substances indispensables à leur croissance du fait de la compétition avec cette dernière. Dans de rares cas, le transport est actif, la plantes mères disposant de moyens pour expulser elle-même à grande distance les graines formées. Le site du Smithsonian nous en propose trois exemples. On parle alors d'autochorie.
Dans le cas ou cette auto-dispersion est impossible, les plantes disposent de trois moyens passifs.
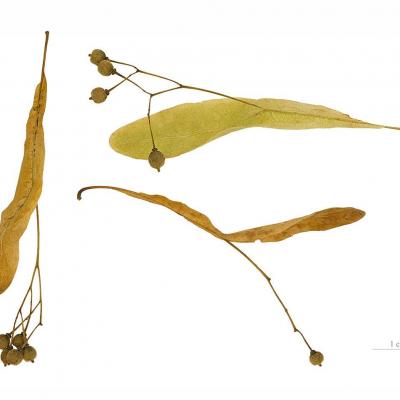
Anémochorie
Le vent est un moyen de dispersion efficace. Dans les deux exemples fournis, celui du tilleul et du pissenlit, on observe des structures qui permettent d'augmenter fortement la portance de la graine. Ainsi, elle pourra, au gré du vent, parcourir des distances importantes et coloniser de nouveaux milieux où elle ne sera plus en compétition avec la plante mère.

Hydrochorie
Les plantes qui vivent dans les milieux humides peuvent utiliser l'eau comme moyen de dispersion. L'un des exemples les plus spectaculaires est celui des noix de coco qui peuvent ainsi traverser des portions d'océan et coloniser des iles qui peuvent être très éloignées du point de départ. Le lotus et le nénuphar font également partie des plantes qui ont recours à l'hydrochorie.

Zoochorie
Les plantes peuvent également s'en remettre aux animaux pour transporter leurs graines. La bardane possède un fruit recouverts de nombreuses barbes qui s'accrochent aisément dans les poils des mammifères. A la faveur d'un nettoyage ou sous l'effet de leur dessication, elles tomberont dans des endroits qu'elles pourront alors coloniser. Dans certains cas, comme celui de la prune, les fruits sont riches en matières organiques (lipides ou glucides) de manière à attirer des animaux qui les consommeront. Une fois ingérées, les graines protégées par une enveloppe dure pourront germer.
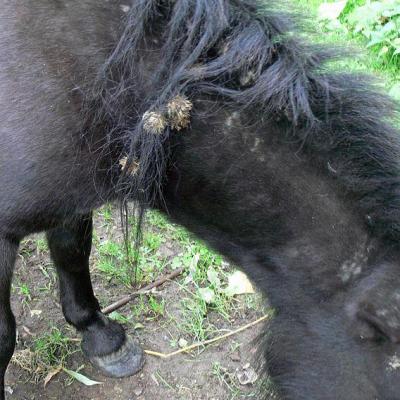
Dans le cas de la zoochorie, on observe fréquemment une coévolution de la plante et de l'animal assurant le transport des graines.
Dans les documents suivants, nous allons traiter de l'exemple d'un cactus, le Melocactus violaceus et d'un lézard resposable de la dissémination des graines, le Tropidurus torquatus.
Le Melocactus violaceus est un cactus d’une dizaine de centimètres de diamètre qui pousse sur les sols sableux des zones désertiques brésiliennes. Il produit des fruits roses au niveau d’un cephalium blanchâtre situé à son sommet.
 Par PierreBraun
Par PierreBraun
Le cactus produit des fruits sucrés et très riches en eau. Ils se forment dans le cephalium et ne sortent qu’à maturité.

Le lézard Tropidurus torquatus est un des rares animaux à pouvoir manger ces fruits.
 Par Albasmalko
Par Albasmalko
La morphologie du lézard lui permet de manger facilement les fruits du cactus : il est assez petit pour se faufiler entre les épines et sa bouche est assez grande pour pouvoir ingérer le fruit. Après digestion, les graines se retrouvent dans les déjections du lézard qui les dépose en moyenne à 3 mètres de la plante mère.

Les fruits du cactus représentent donc une importante ressource alimentaire et hydrique pour les lézards. Sans ces fruits, la survie du lézard est compromise. Seuls les lézards ayant la capacité de se nourrir de ces fruits ont eu la possibilité de transmettre leur patrimoine génétique, induisant ainsi une sélection des individus les plus adaptés à ce milieu. On a donc bien affaire à un phénomène d'évolution.
Une équipe de chercheurs a mesuré le rythme de sortie des fruits de 118 cactus pendant une journée et l’a mis en parallèle avec le nombre de lézards présents autour des plantes.
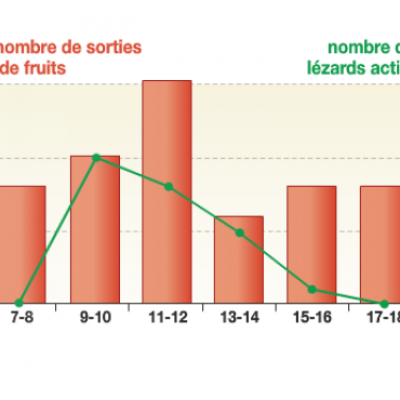
On observe dans le graphique exprimant les résultats de cette étude que la sortie des fruits coïncide avec les horaires où les lézards sont les plus actifs, soit entre 9 heure et midi. La maturation des fruits est donc coordonnée au rythme de vie des lézards. L'écueil serait d'y voir une volonté du cactus pour que ses fruits puissent être mangés et ses graines disséminées par les lézards. En réalité, seuls les cactus dont la période de maturation des fruits est coordonnée au cycle de vie des lézards voient leurs graines disséminées. Nous sommes donc bien en présence d'un phénomène d'évolution.
Pour estimer le pouvoir germinatif des graines digérées, des chercheurs ont récupéré et planté des graines mangées par des lézards. Ils ont suivi le taux de germination de ces graines au cours du temps en comparaison avec des graines n’ayant pas transit é par le système digestif d’un lézard.
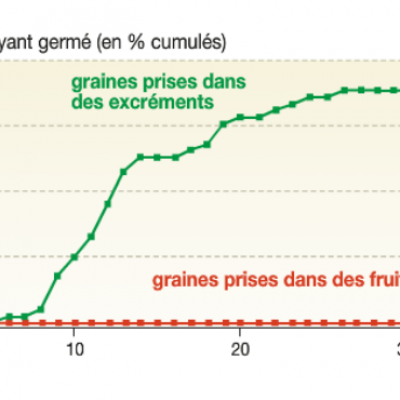
On observe dans ce graphique que seules les graines ayant transité dans le système digestif des lézards peuvent germer, ce qui indique une coévolution entre le cactus et le lézard. En effet, au cours du temps, seules les graines pouvant résister au système digestif du lézard ont pu entraîner le développement de la plante. Par conséquent, les graines qui peuvent germer doivent passer dans le système digestif pour pouvoir germer. Encore une fois, nous sommes bien en présence d'un phénomène d'évolution.
Il n'est donc pas inexact de mentionner un phénomène de coévolution entre le lézard et le cactus. Seuls les lézards capables de se nourrir des fruits du cactus sont sélectionnés, seuls les cactus dont les fruits sont matures aux périodes d'activité et dont les graines germent après passage dans le système digestifs sont sélectionnés.
A mardi prochain pour la suite.
Date de dernière mise à jour : 30/04/2020
Ajouter un commentaire